
Tubuh manusia tidak mungkin terhindar dari lingkungan yang mengandung mikroba patogen di sekelilingnya. Mikroba tersebut dapat menimbulkan penyakit infeksi pada manusia. Mikroba patogen yang ada bersifat poligenik dan kompleks. Oleh karena itu respons imun tubuh manusia terhadap berbagai macam mikroba patogen juga berbeda. Umumnya gambaran biologik spesifik mikroba menentukan mekanisme imun mana yang berperan untuk proteksi. Begitu juga respon imun terhadap bakteri khususnya bakteri ekstraselular atau bakteri intraselular mempunyai karakteristik tertentu pula
Tubuh manusia akan selalu terancam oleh paparan bakteri, virus, parasit, radiasi matahari, dan polusi. Stres emosional atau fisiologis dari kejadian ini adalah tantangan lain untuk mempertahankan tubuh yang sehat. Biasanya kita dilindungi oleh sistem pertahanan tubuh, sistem kekebalan tubuh, terutama makrofag, dan cukup lengkap kebutuhan gizi untuk menjaga kesehatan. Kelebihan tantangan negatif, bagaimanapun, dapat menekan sistem pertahanan tubuh, sistem kekebalan tubuh, dan mengakibatkan berbagai penyakit fatal.
Penerapan kedokteran klinis saat ini adalah untuk mengobati penyakit saja. Infeksi bakteri dilawan dengan antibiotik, infeksi virus dengan antivirus dan infeksi parasit dengan antiparasit terbatas obat-obatan yang tersedia. Sistem pertahanan tubuh, sistem kekebalan tubuh, depresi disebabkan oleh stres emosional diobati dengan antidepresan atau obat penenang. Kekebalan depresi disebabkan oleh kekurangan gizi jarang diobati sama sekali, bahkan jika diakui, dan kemudian oleh saran untuk mengkonsumsi makanan yang lebih sehat.
Imunitas atau kekebalan adalah sistem mekanisme pada organisme yang melindungi tubuh terhadap pengaruh biologis luar dengan mengidentifikasi dan membunuh patogen serta sel tumor. Sistem ini mendeteksi berbagai macam pengaruh biologis luar yang luas, organisme akan melindungi tubuh dari infeksi, bakteri, virus sampai cacing parasit, serta menghancurkan zat-zat asing lain dan memusnahkan mereka dari sel organisme yang sehat dan jaringan agar tetap dapat berfungsi seperti biasa. Deteksi sistem ini sulit karena adaptasi patogen dan memiliki cara baru agar dapat menginfeksi organisme.
Untuk selamat dari tantangan ini, beberapa mekanisme telah berevolusi yang menetralisir patogen. Bahkan organisme uniselular seperti bakteri dimusnahkan oleh sistem enzim yang melindungi terhadap infeksi virus. Mekanisme imun lainnya yang berevolusi pada eukariota kuno dan tetap pada keturunan modern, seperti tanaman, ikan, reptil dan serangga. Mekanisme tersebut termasuk peptida antimikrobial yang disebut defensin, fagositosis, dan sistem komplemen. Mekanisme yang lebih berpengalaman berkembang secara relatif baru-baru ini, dengan adanya evolusi vertebrata. Imunitas vertebrata seperti manusia berisi banyak jenis protein, sel, organ tubuh dan jaringan yang berinteraksi pada jaringan yang rumit dan dinamin. Sebagai bagian dari respon imun yang lebih kompleks ini, sistem vertebrata mengadaptasi untuk mengakui patogen khusus secara lebih efektif. Proses adaptasi membuat memori imunologis dan membuat perlindungan yang lebih efektif selama pertemuan di masa depan dengan patogen tersebut. Proses imunitas yang diterima adalah basis dari vaksinasi.
Respons pejamu yang terjadi juga tergantung dari jumlah mikroba yang masuk. Mekanisme pertahanan tubuh dalam mengatasi agen yang berbahaya meliputi
- Pertahanan fisik dan kimiawi, seperti kulit, sekresi asam lemak dan asam laktat melalui kelenjar keringat, sekresi lendir, pergerakan silia, sekresi air mata, air liur, urin, asam lambung serta lisosom dalam air mata
- Simbiosis dengan bakteri flora normal yang memproduksi zat yang dapat mencegah invasi mikroorganisme
- Innate immunity (mekanisme non-spesifik), seperti sel polimorfonuklear (PMN) dan makrofag, aktivasi komplemen, sel mast, protein fase akut, interferon, sel NK (natural killer) dan mediator eosinofil
- Imunitas spesifik, yang terdiri dari imunitas humoral dan seluler. Secara umum pengontrolan infeksi intraselular seperti infeksi virus, protozoa, jamur dan beberapa bakteri intraselular fakultatif terutama membutuhkan imunitas yang diperani oleh sel yang dinamakan imunitas selular, sedangkan bakteri ekstraselular dan toksin membutuhkan imunitas yang diperani oleh antibodi yang dinamakan imunitas humoral. Secara keseluruhan pertahanan imunologik dan nonimunologik (nonspesifik) bertanggung jawab bersama dalam pengontrolan terjadinya penyakit infeksi.
Keberhasilan patogen bergantung pada kemampuannya untuk menghindar dari respon imun. Patogen telah mengembangkan beberapa metode yang menyebabkan mereka dapat menginfeksi sementara patogen menghindari kehancuran akibat sistem imun. Bakteri sering menembus perisai fisik dengan mengeluarkan enzim yang mendalami isi perisai, contohnya dengan menggunakan sistem tipe II sekresi. Sebagai kemungkinan, patogen dapat menggunakan sistem tipe III sekresi. Mereka dapat memasukan tuba palsu pada sel, yang menyediakan saluran langsung untuk protein agar dapat bergerak dari patogen ke pemilik tubuh; protein yang dikirim melalui tuba sering digunakan untuk mematikan pertahanan.
Strategi menghindari digunakan oleh beberapa patogen untuk mengelakan sistem imun bawaan adalah replikasi intraselular (juga disebut patogenesis intraselular). Disini, patogen mengeluarkan mayoritas lingkaran hidupnya kedalam sel yang dilindungi dari kontak langsung dengan sel imun, antibodi dan komplemen. Beberapa contoh patogen intraselular termasuk virus, racun makanan, bakteri Salmonella dan parasit eukariot yang menyebabkan malaria (Plasmodium falciparum) dan leismaniasis (Leishmania spp.). Bakteri lain, seperti Mycobacterium tuberculosis, hidup didalam kapsul protektif yang mencegah lisis oleh komplemen. Banyak patogen mengeluarkan senyawa yang mengurangi respon imun atau mengarahkan respon imun ke arah yang salah. Beberapa bakteri membentuk biofilm untuk melindungi diri mereka dari sel dan protein sistem imun. Biofilm ada pada banyak infeksi yang berhasil, seperti Pseudomonas aeruginosa kronik dan Burkholderia cenocepacia karakteristik infeksi sistik fibrosis. Bakteri lain menghasilkan protein permukaan yang melilit pada antibodi, mengubah mereka menjadi tidak efektif; contoh termasuk Streptococcus (protein G), Staphylococcus aureus (protein A), dan Peptostreptococcus magnus (protein L).

Bakteri, dari kata Latin bacterium (jamak, bacteria), adalah kelompok terbanyak dari organisme hidup. Mereka sangatlah kecil (mikroskopik) dan kebanyakan uniselular (bersel tunggal), dengan struktur sel yang relatif sederhana tanpa nukleus/inti sel, cytoskeleton, dan organel lain seperti mitokondria dan kloroplas. Struktur sel mereka dijelaskan lebih lanjut dalam artikel mengenai prokariota, karena bakteri merupakan prokariota, untuk membedakan mereka dengan organisme yang memiliki sel lebih kompleks, disebut eukariota. Istilah “bakteri” telah diterapkan untuk semua prokariota atau untuk kelompok besar mereka, tergantung pada gagasan mengenai hubungan mereka.
Bakteri adalah yang paling berkelimpahan dari semua organisme. Mereka tersebar (berada di mana-mana) di tanah, air, dan sebagai simbiosis dari organisme lain. Banyak patogen merupakan bakteri. Kebanyakan dari mereka kecil, biasanya hanya berukuran 0,5-5 μm, meski ada jenis dapat menjangkau 0,3 mm dalam diameter (Thiomargarita). Mereka umumnya memiliki dinding sel, seperti sel tumbuhan dan jamur, tetapi dengan komposisi sangat berbeda (peptidoglikan). Banyak yang bergerak menggunakan flagela, yang berbeda dalam strukturnya dari flagela kelompok lain.
 \
\SPECIFIC ATTACHMENTS OF BACTERIA TO HOST CELL OR TISSUE SURFACES
Adhesin | Receptor | Attachment site | Disease | |
Streptococcus pyogenes | Protein F | Amino terminus of fibronectin | Pharyngeal epithelium | Sore throat |
Streptococcus mutans | Glycosyl transferase | Salivary glycoprotein | Pellicle of tooth | Dental caries |
Streptococcus salivarius | Lipoteichoic acid | Unknown | Buccal epithelium of tongue | None |
Streptococcus pneumoniae | Cell-bound protein | N-acetylhexosamine-galactose disaccharide | Mucosal epithelium | pneumonia |
Staphylococcus aureus | Cell-bound protein | Amino terminus of fibronectin | Mucosal epithelium | Various |
Neisseria gonorrhoeae | Type IV pili (N-methylphenyl- alanine pili) | Glucosamine-galactose carbohydrate | Urethral/cervical epithelium | Gonorrhea |
Enterotoxigenic E. coli | Type-I fimbriae | Species-specific carbohydrate(s) | Intestinal epithelium | Diarrhea |
Uropathogenic E. coli | Type I fimbriae | Complex carbohydrate | Urethral epithelium | Urethritis |
Uropathogenic E. coli | P-pili (pap) | Globobiose linked to ceramide lipid | Upper urinary tract | Pyelonephritis |
Bordetella pertussis | Fimbriae (“filamentous hemagglutinin”) | Galactose on sulfated glycolipids | Respiratory epithelium | Whooping cough |
Vibrio cholerae | N-methylphenylalanine pili | Fucose and mannose carbohydrate | Intestinal epithelium | Cholera |
Treponema pallidum | Peptide in outer membrane | Surface protein (fibronectin) | Mucosal epithelium | Syphilis |
Mycoplasma | Membrane protein | Sialic acid | Respiratory epithelium | Pneumonia |
Chlamydia | Unknown | Sialic acid | Conjunctival or urethral epithelium |
Strategi pertahanan bakteri
Bakteri ekstraseluler adalah bakteri yang dapat bereplikasi di luar sel, di dalam sirkulasi, di jaringan ikat ekstraseluler, dan di berbagai jaringan. Berbagai jenis bakteri yang termasuk golongan bakteri ekstraseluler telah disebutkan pada bab sebelumnya. Bakteri ekstraseluler biasanya mudah dihancurkan oleh sel fagosit. Pada keadaan tertentu bakteri ekstraseluler tidak dapat dihancurkan oleh sel fagosit karena adanya sintesis kapsul antifagosit, yaitu kapsul luar (outer capsule) yang mengakibatkan adesi yang tidak baik antara sel fagosit dengan bakteri, seperti pada infeksi bakteri berkapsul Streptococcus pneumoniae atau Haemophylus influenzae. Selain itu, kapsul tersebut melindungi molekul karbohidrat pada permukaan bakteri yang seharusnya dapat dikenali oleh reseptor fagosit. Dengan adanya kapsul ini, akses fagosit dan deposisi C3b pada dinding sel bakteri dapat dihambat. Beberapa organisme lain mengeluarkan eksotoksin yang meracuni leukosit. Strategi lainnya adalah dengan pengikatan bakteri ke permukaan sel non fagosit sehingga memperoleh perlindungan dari fungsi fagosit .
Sel normal dalam tubuh mempunyai protein regulator yang melindungi dari kerusakan oleh komplemen, seperti CR1, MCP dan DAF, yang menyebabkan pemecahan C3 konvertase. Beberapa bakteri tidak mempunyai regulator tersebut, sehingga akan mengaktifkan jalur alternatif komplemen melalui stabilisasi C3b3b konvertase pada permukaan sel bakteri. Dengan adanya kapsul bakteri akan menyebabkan aktivasi dan stabilisasi komplemen yang buruk.
Beberapa bakteri juga dapat mempercepat pemecahan komplemen melalui aksi produk mikrobial yang mengikat atau menghambat kerja regulator aktivasi komplemen. Bahkan beberapa spesies dapat menghindari lisis dengan cara mengalihkan lokasi aktivasi komplemen melalui sekresi protein umpan (decoy protein) atau posisi permukaan bakteri yang jauh dari membran sel. Beberapa organisme Gram positif mempunyai lapisan peptidoglikan tebal yang menghambat insersi komplek serangan membran C5b-9 pada membran sel bakteri .
Bakteri enterik Gram negatif pada usus mempengaruhi aktivitas makrofag termasuk menginduksi apoptosis, meningkatkan produksi IL-1, mencegah fusi fagosom-lisosom dan mempengaruhi sitoskleton aktin. Strategi berupa variasi antigenik juga dimiliki oleh beberapa bakteri, seperti variasi lipoprotein permukaan, variasi enzim yang terlibat dalam sintesis struktur permukaan dan variasi antigenik pili.Keadaan sistem imun yang dapat menyebabkan bakteri ekstraseluler sulit dihancurkan adalah gangguan pada mekanisme fagositik karena defisiensi sel fagositik (neutropenia) atau kualitas respons imun yang kurang (penyakit granulomatosa kronik).
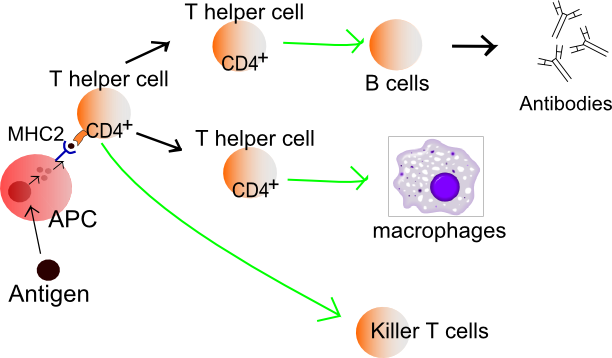

Mekanisme pertahanan bakteri ekstraseluler.

Invasin | Bacteria Involved | Activity |
| Hyaluronidase | Streptococci, staphylococci and clostridia | Degrades hyaluronic of connective tissue |
| Collagenase | Clostridium species | Dissolves collagen framework of muscles |
| Neuraminidase | Vibrio cholerae and Shigella dysenteriae | Degrades neuraminic acid of intestinal mucosa |
| Coagulase | Staphylococcus aureus | Converts fibrinogen to fibrin which causes clotting |
| Kinases | Staphylococci and streptococci | Converts plasminogen to plasmin which digests fibrin |
| Leukocidin | Staphylococcus aureus | Disrupts neutrophil membranes and causes discharge of lysosomal granules |
| Streptolysin | Streptococcus pyogenes | Repels phagocytes and disrupts phagocyte membrane and causes discharge of lysosomal granules |
| Hemolysins | Streptococci, staphylococci and clostridia | Phospholipases or lecithinases that destroy red blood cells (and other cells) by lysis |
| Lecithinases | Clostridium perfringens | Destroy lecithin in cell membranes |
| Phospholipases | Clostridium perfringens | Destroy phospholipids in cell membrane |
| Anthrax EF | Bacillus anthracis | One component (EF) is an adenylate cyclase which causes increased levels of intracellular cyclic AMP |
| Pertussis AC | Bordetella pertussis | One toxin component is an adenylate cyclase that acts locally producing an increase in intracellular cyclic AMP |
Respons imun terhadap bakteri ekstraseluler bertujuan untuk menetralkan efek toksin dan mengeliminasi bakteri. Respons imun alamiah terutama melalui fagositosis oleh neutrofil, monosit serta makrofag jaringan. Lipopolisakarida dalam dinding bakteri Gram negatif dapat mengaktivasi komplemen jalur alternatif tanpa adanya antibodi. Hasil aktivasi ini adalah C3b yang mempunyai efek opsonisasi, lisis bakteri melalui serangan kompleks membran dan respons inflamasi akibat pengumpulan serta aktivasi leukosit. Endotoksin juga merangsang makrofag dan sel lain seperti endotel vaskular untuk memproduksi sitokin seperti TNF, IL-1, IL-6 dan IL-8. Sitokin akan menginduksi adesi neutrofil dan monosit pada endotel vaskular pada tempat infeksi, diikuti dengan migrasi, akumulasi lokal serta aktivasi sel inflamasi. Kerusakan jaringan yang terjadi adalah akibat efek samping mekanisme pertahanan untuk eliminasi bakteri. Sitokin juga merangsang demam dan sintesis protein fase akut.
Netralisasi toksin
Infeksi bakteri Gram negatif dapat menyebabkan pengeluaran endotoksin yang akan menstimulasi makrofag. Stimulasi yang berlebihan terhadap makrofag akan menghasilkan sejumlah sitokin seperti IL-1, IL-6 dan TNF. Proses ini akan memacu terjadinya reaksi peradangan yang menyebabkan kerusakan sel, hipotensi, aktivasi sistem koagulasi, gagal organ multipel dan berakhir dengan kematian. Antibodi yang mengandung reseptor sitokin dan antagonisnya, berperan dalam menghilangkan sejumlah sitokin dalam sirkulasi dan mencegah sitokin berikatan pada sel target.
Antibodi yang beredar dalam sirkulasi akan menetralisasi molekul antifagositik dan eksotoksin lainnya yang diproduksi bakteri. Mekanisme netralisasi antibodi terhadap bakteri terjadi melalui dua cara. Pertama, melalui kombinasi antibodi di dekat lokasi biologi aktif infeksi yaitu secara langsung menghambat reaksi toksin dengan sel target. Kedua, melalui kombinasi antibodi yang terletak jauh dari lokasi biologi aktif infeksi yaitu dengan mengubah konformasi alosterik toksin agar tidak dapat bereaksi dengan sel target. Dengan ikatan kompleks bersama antibodi, toksin tidak dapat berdifusi sehingga rawan terhadap fagositosis, terutama bila ukuran kompleks membesar karena deposisi komplemen pada permukaan bakteri akan semakin bertambah.
Opsonisasi
Opsonisasi adalah pelapisan antigen oleh antibodi, komplemen, fibronektin, yang berfungsi untuk memudahkan fagositosis. Opsonisasi ada dua yaitu opsonisasi yang tidak tergantung antibodi dan yang ditingkatkan oleh antibodi.
Pada opsonisasi yang tidak tergantung antibodi, protein pengikat manose dapat terikat pada manose terminal pada permukaan bakteri, dan akan mengaktifkan C1r dan C1s serta berikatan dengan C1q. Proses tersebut akan mengaktivasi komplemen pada jalur klasik yang dapat berperan sebagai opsonin dan memperantarai fagositosis. Lipopolisakarida (LPS) merupakan endotoksin yang penting pada bakteri Gram negatif. Sel ini dapat dikenal oleh tiga kelas molekul reseptor. Sedangkan opsonisasi yang ditingkatkan oleh antibodi adalah bakteri yang resisten terhadap proses fagositosis akan tertarik pada sel PMN dan makrofag bila telah diopsonisasi oleh antibodi.
Dalam opsonisasi terdapat sinergisme antara antibodi dan komplemen yang diperantarai oleh reseptor yang mempunyai afinitas kuat untuk IgG dan C3b pada permukaan fagosit, sehingga meningkatkan pengikatan di fagosit. Efek augmentasi dari komplemen berasal dari molekul IgG yang dapat mengikat banyak molekul C3b, sehingga meningkatkan jumlah hubungan ke makrofag (bonus effect of multivalency). Meskipun IgM tidak terikat secara spesifik pada makrofag, namun merangsang adesi melalui pengikatan komplemen.
Antibodi akan menginisiasi aksi berantai komplemen sehingga lisozim serum dapat masuk ke dalam lapisan peptidoglikan bakteri dan menyebabkan kematian sel. Aktivasi komplemen melalui penggabungan dengan antibodi dan bakteri juga menghasilkan anfilaktoksin C3a dan C5a yang berujung pada transudasi luas dari komponen serum, termasuk antibodi yang lebih banyak, dan juga faktor kemotaktik terhadap neutrofil untuk membantu fagositosis.
Sel PMN merupakan fagosit yang predominan dalam sirkulasi dan selalu tiba di lokasi infeksi lebih cepat dari sel lain, karena sel PMN tertarik oleh sinyal kemotaktik yang dikeluarkan oleh bakteri, sel PMN lain, komplemen atau makrofag lain, yang lebih dahulu tiba di tempat infeksi. Sel PMN sangat peka terhadap semua faktor kemotaktik.
Sel PMN yang telah mengalami kemotaktik selanjutnya akan melakukan adesi pada dinding sel bakteri, endotel maupun jaringan yang terinfeksi. Kemampuan adesi PMN pada permukaan sel bakteri akan bertambah kuat karena sinyal yang terbentuk pada proses adesi ini akan merangsang ekspresi Fc dan komplemen pada permukaan sel. Sel PMN juga akan melakukan proses diapedesis agar dapat menjangkau bakteri yang telah menginfeksi.
Proses penelanan bakteri oleh fagosit diawali dengan pembentukan tonjolan pseudopodia yang berbentuk kantong fagosom untuk mengelilingi bakteri, sehingga bakteri akan terperangkap di dalamnya, selanjutnya partikel granular di dalam fagosom akan mengeluarkan berbagai enzim dan protein untuk merusak dan menghancurkan bakteri tersebut.
Mekanisme pemusnahan bakteri oleh enzim ini dapat melalui proses oksidasi maupun nonoksidasi, tergantung pada jenis bakteri dan status metabolik pada saat itu. Oksidasi dapat berlangsung dengan atau tanpa mieloperoksidase. Proses oksidasi dengan mieloperoksidase terjadi melalui ikatan H2O2 dengan Fe yang terdapat pada mieloperoksidase. Proses ini menghasilkan komplek enzim-subtrat dengan daya oksidasi tinggi dan sangat toksik terhadap bakteri, yaitu asam hipoklorat (HOCl).
Proses oksidasi tanpa mieloperoksidase berdasarkan ikatan H2O2 dengan superoksida dan radikal hidroksil namun daya oksidasinya rendah. Proses nonoksidasi berlangsung dengan perantaraan berbagai protein dalam fagosom yaitu flavoprotein, sitokrom-b, laktoferin, lisozim, kaptensin G dan difensin. Pada proses pemusnahan bakteri, pH dalam sel fagosit dapat menjadi alkalis. Hal ini terjadi karena protein yang bermuatan positif dalam pH yang alkalis bersifat sangat toksik dan dapat merusak lapisan lemak dinding bakteri Gram negatif. Selain itu, bakteri juga dapat terbunuh pada saat pH dalam fagosom menjadi asam karena aktivitas lisozim. Melalui proses ini PMN memproduksi antibakteri yang dapat berperan sebagai antibiotika alami (natural antibiotics).
Sistem imun sekretori
Permukaan mukosa usus mempunyai mekanisme pertahanan spesifik antigen dan nonspesifik. Mekanisme nonspesifik terdiri dari peptida antimikrobial yang diproduksi oleh neutrofil, makrofag dan epitel mukosa. Peptida ini akan menyebabkan lisis bakteri melalui disrupsi pada permukaan membran. Imunitas spesifik diperantarai oleh IgA sekretori dan IgM, dengan dominasi IgA1 pada usus bagian awal dan IgA2 pada usus besar. Antibodi IgA mempunyai fungsi proteksi dengan cara melapisi (coating) virus dan bakteri dan mencegah adesi pada sel epitel di membran mukosa. Reseptor Fc dari kelas Ig ini mempunyai afinitas tinggi terhadap neutrofil dan makrofag dalam proses fagositosis. Apabila agen infeksi berhasil melewati barier IgA, maka lini pertahanan berikutnya adalah IgE. Adanya kontak antigen dengan IgE akan menyebabkan pelepasan mediator yang menarik agen respons imun dan menghasilkan reaksi inflamasi akut. Adanya peningkatan permeabilitas vaskular yang disebabkan oleh histamin akan menyebabkan transudasi IgG dan komplemen, sedangkan faktor kemotaktik terhadap neutrofil dan eosinofil akan menarik sel efektor yang diperlukan untuk mengatasi organisme penyebab infeksi yang telah dilapisi oleh IgG spesifik dan C3b. Penyatuan kompleks antibodi-komplemen pada makrofag akan menghasilkan faktor yang memperkuat permeabilitas vaskular dan proses kemotaktik .
Apabila organisme yang diopsonisasi terlalu besar untuk difagosit, maka fagosit dapat mengatasi organisme tersebut melalui mekanisme ekstraseluler, yaitu Antibody-Dependent Cellular Cytotoxicity (ADCC).
INFEKSI BAKTERI INTRASELULER
Strategi pertahanan bakteri

Bakteri intraseluler terbagi atas dua jenis, yaitu bakteri intraseluler fakultatif dan obligat. Bakteri intraseluler fakultatif adalah bakteri yang mudah difagositosis tetapi tidak dapat dihancurkan oleh sistem fagositosis. Bakteri intraseluler obligat adalah bakteri yang hanya dapat hidup dan berkembang biak di dalam sel hospes. Hal ini dapat terjadi karena bakteri tidak dapat dijangkau oleh antibodi dalam sirkulasi, sehingga mekanisme respons imun terhadap bakteri intraseluler juga berbeda dibandingkan dengan bakteri ekstraseluler. Beberapa jenis bakteri seperti basil tuberkel dan leprosi, dan organisme Listeria dan Brucella menghindari perlawanan sistem imun dengan cara hidup intraseluler dalam makrofag, biasanya fagosit mononuklear, karena sel tersebut mempunyai mobilitas tinggi dalam tubuh. Masuknya bakteri dimulai dengan ambilan fagosit setelah bakteri mengalami opsonisasi. Namun setelah di dalam makrofag, bakteri tersebut melakukan perubahan mekanisme pertahanan.
Bakteri intraseluler memiliki kemampuan mempertahankan diri melalui tiga mekanisme, yaitu 1) hambatan fusi lisosom pada vakuola yang berisi bakteri, 2) lipid mikobakterial seperti lipoarabinomanan menghalangi pembentukan ROI (reactive oxygen intermediate) seperti anion superoksida, radikal hidroksil dan hidrogen peroksida dan terjadinya respiratory burst, 3) menghindari perangkap fagosom dengan menggunakan lisin sehingga tetap hidup bebas dalam sitoplasma makrofag dan terbebas dari proses pemusnahan selanjutnya (Gambar 13-4).


Mekanisme pertahanan tubuh

Pertahanan oleh diperantarai sel T (Celluar Mediated Immunity, CMI) sangat penting dalam mengatasi organisme intraseluler. Sel T CD4 akan berikatan dengan partikel antigen yang dipresentasikan melalui MHC II pada permukaan makrofag yang terinfeksi bakteri intraseluler. Sel T helper (Th1) ini akan mengeluarkan sitokin IFN γ yang akan mengaktivasi makrofag dan membunuh organisme intraseluler, terutama melalui pembentukan oksigen reaktif intermediat (ROI) dan nitrit oxide (NO). Selanjutnya makrofag tersebut akan mengeluarkan lebih banyak substansi yang berperan dalam reaksi inflamasi kronik. Selain itu juga terjadi lisis sel yang diperantarai oleh sel T CD8.
Beberapa bakteri ada yang resisten sehingga menimbulkan stimulasi antigen yang kronik. Keadaan ini menimbulkan pengumpulan lokal makrofag yang terkativasi yang membentuk granuloma sekeliling mikroorganisme untuk mencegah penyebaran. Hal ini dapat berlanjut pada nekrosis jaringan dan fibrosis yang luas yang menyebabkan gangguan fungsi. Oleh karena itu, kerusakan jaringan terutama disebabkan oleh respons imun terhadap infeksi bakteri intraseluler.
Daftar Pustaka
- Hardegree MC, Tu AT (eds): Handbook of Natural Toxins. Vol.4: Bacterial Toxins. Marcel Dekker, New York, 1988
- Iglewski BH, Clark VL (eds): Molecular Basis of Bacterial Pathogenesis. Vol. XI of The Bacteria: A Treatise on Structure and Function. Academic Press, Orlando, FL, 1990
- Buku Ajar Alergi Imunologi. Ikatan Dokter Anak Indonesia edisi 2.
- Luderitz O, Galanos C: Endotoxins of gram-negative bacteria. p.307. In Dorner F, Drews J (eds): Pharmacology of Bacterial Toxins. International Encyclopedia of Pharmacology and Therapeutics, Section 119. Pergamon, Elmsford, NY, 1986
- Mims CA: The Pathogenesis of Infectious Disease. Academic Press, London, 1976
- Payne SM: Iron and virulence in the family Enterobacteriaceae. Crit Rev Microbiol 16:81, 1988
- Sack RB: Human diarrheal disease caused by enterotoxigenic Escherichia coli. Annu Rev Microbiol 29:333, 1975
- Salyers, AA, Whitt DD: Bacterial Pathogenesis – A Molecular Approach ASM Press, 1994
- Smith H: Microbial surfaces in relation to pathogenicity. Bacteriol Rev 41:475, 1977
- Smith H, Turner JJ (eds): The Molecular Basis of Pathogenicity. Verlag Chemie, Deerfield Beach, FL, 1980
- Weinberg ED: Iron withholding: a defense against infection and neoplasia. Physiol Rev 64:65, 1984
- Eisenstein TK, Actor P, Friedman H: Host Defenses to Intracellular Pathogens. Plenum Publishing Co, New York, 1983
- Finlay BB, Falkow S: Common themes in microbial pathogenicity. Microbiol Rev 53:210, 1989
- Foster TJ: Plasmid-determined resistance to antimicrobial drugs and toxic metal ions in bacteria. Microbiol Rev 47:361, 1983
Tidak ada komentar:
Posting Komentar